Research Insight
Transposable Elements in Zea: Their Role in Genetic Diversity and Evolution 
 Author
Author  Correspondence author
Correspondence author
Maize Genomics and Genetics, 2024, Vol. 15, No. 3 doi: 10.5376/mgg.2024.15.0013
Received: 14 Apr., 2024 Accepted: 20 May, 2024 Published: 02 Jun., 2024
Yang S.M., 2024, Transposable elements in zea: their role in genetic diversity and evolution, Maize Genomics and Genetics, 15(3): 123-135 (doi: 10.5376/mgg.2024.15.0013)
Transposable elements (TEs) are significant contributors to genetic diversity and evolutionary processes in Zea mays (maize). These mobile genetic elements can move within the genome, inducing mutations, structural variations, and changes in gene expression, which collectively enhance genetic variability and adaptability. TEs are maintained in a delicate balance within the genome, as they can be both deleterious and beneficial. In maize, TEs have been shown to play crucial roles in genome evolution, including the generation of allelic diversity and the regulation of gene expression. The maize genome is particularly rich in TEs, with recent advancements in annotation methods revealing a higher abundance and diversity of TEs than previously recognized. Moreover, the interplay between TEs and epigenetic mechanisms, such as RNA N6-methyladenosine modification, further underscores their role in the adaptive evolution of maize. This study synthesizes current knowledge on the impact of TEs on the genetic diversity and evolutionary dynamics of maize, highlighting their dual role as both genomic parasites and symbionts. The findings underscore the importance of TEs in shaping the maize genome and their potential in driving adaptive responses to environmental challenges.
1 Introduction
Transposable elements (TEs) are mobile DNA sequences that can move from one location to another within a genome. They are ubiquitous across all forms of life, from bacteria to humans, and play a significant role in shaping genomic architecture and function. TEs can be classified into two major types: transposons, which move directly via a "cut-and-paste" mechanism, and retrotransposons, which move indirectly through an RNA intermediate (Fedoroff, 2012; Romano and Fanti, 2022). The activity of TEs can lead to various genomic alterations, including gene disruptions, chromosomal rearrangements, and the creation of new regulatory sequences (Chénais et al., 2011; Bennetzen and Wang, 2014). While TEs are often considered genomic parasites due to their potential to cause deleterious mutations, they also contribute to genetic diversity and evolutionary innovation by providing new genetic material and regulatory elements (Rebollo et al., 2012; Sundaram and Wysocka, 2020; Romano and Fanti, 2022).
Zea mays, commonly known as maize, is a model organism for studying TEs due to its large and complex genome, which is rich in TEs. The maize genome is composed of a significant proportion of TEs, making it an ideal system to explore the impact of these elements on genome structure, function, and evolution (Fedoroff, 2012; Hirsch and Springer, 2017). The study of TEs in maize has historical significance, as the first discovery of TEs was made by Barbara McClintock in maize plants, leading to groundbreaking insights into genetic regulation and genome dynamics (Fedoroff, 2012). Understanding the role of TEs in maize is crucial for several reasons: it can provide insights into the mechanisms of genome evolution, reveal how TEs contribute to genetic diversity and adaptation, and inform breeding strategies for crop improvement (Vicient and Casacuberta, 2017; Platt et al., 2018). Moreover, the activity of TEs in maize can influence gene expression and regulatory networks, thereby affecting phenotypic traits and responses to environmental stresses.
This study is to provide a comprehensive overview of the role of transposable elements in the genetic diversity and evolution of Zea mays. This study will cover the following key areas: the classification and mechanisms of TEs, the historical and contemporary significance of TEs in maize, the impact of TEs on genome structure and function, and the evolutionary implications of TE activity. By synthesizing findings from recent research, this study aims to highlight the dual nature of TEs as both genomic parasites and beneficial elements, and to elucidate their contributions to the genetic and phenotypic diversity of maize. Through this study hope to advance the understanding of TEs in maize and their broader implications for plant genomics and evolution.
2 Types of Transposable Elements in Zea
2.1 DNA transposons
DNA transposons, also known as Class II transposable elements, move within the genome through a "cut-and-paste" mechanism. This process involves the excision of the transposon from one location and its reinsertion at another site within the genome. DNA transposons are characterized by terminal inverted repeats (TIRs) and the presence of a transposase enzyme that facilitates their movement. In maize, DNA transposons have been shown to play significant roles in genome evolution by inducing structural variations and generating allelic diversity. For instance, the Activator (Ac) and Dissociation (Ds) elements are well-studied DNA transposons in maize that can cause complex chromosomal rearrangements, leading to variations in gene expression and phenotypic diversity (Fedoroff, 2012; Su et al., 2019; Sharma and Peterson, 2022).
2.2 Retrotransposons
Retrotransposons, or Class I transposable elements, move within the genome via an RNA intermediate. This "copy-and-paste" mechanism involves the transcription of the retrotransposon into RNA, which is then reverse-transcribed into DNA and inserted at a new location. Retrotransposons are further classified into long terminal repeat (LTR) and non-LTR retrotransposons. In maize, retrotransposons constitute a significant portion of the genome, contributing to its large size and complexity. They are often associated with regulatory elements that can influence gene expression. For example, studies have shown that retrotransposon-derived sequences can act as cis-regulatory elements, affecting the transcriptional regulation of nearby genes (Zhao et al., 2018; Stitzer et al., 2019; Noshay et al., 2020).
2.3 Specific TEs identified in maize (e.g., Ac/Ds, Spm/En)
In maize, several specific transposable elements have been identified and extensively studied for their roles in genetic diversity and evolution. The Ac/Ds system is one of the most well-known DNA transposon systems in maize. The Ac element encodes a transposase that can mobilize both Ac and Ds elements. The movement of these elements can cause mutations, gene disruptions, and chromosomal rearrangements, contributing to genetic diversity (Fedoroff, 2012; Wang et al., 2020; Sharma and Peterson, 2022). Another significant DNA transposon system in maize is the Suppressor-mutator (Spm) or Enhancer (En) system. Similar to Ac/Ds, the Spm/En elements can induce mutations and alter gene expression, further adding to the genetic variability within maize populations (Fedoroff, 2012; Sharma and Peterson, 2022).
Retrotransposons also play a crucial role in maize genome dynamics. For instance, the long terminal repeat (LTR) retrotransposons are abundant in the maize genome and have been implicated in the regulation of gene expression. These elements can insert near genes and provide regulatory sequences that enhance or repress gene activity. The presence of retrotransposon-derived cis-regulatory elements has been shown to significantly impact the transcriptional landscape of maize, highlighting their importance in genome evolution (Zhao et al., 2018; Stitzer et al., 2019; Noshay et al., 2020).
The diverse array of transposable elements in maize, including both DNA transposons and retrotransposons, underscores their critical role in shaping the genetic architecture and evolutionary trajectory of this species. By generating genetic diversity and influencing gene expression, transposable elements contribute to the adaptability and phenotypic plasticity of maize, making them a key focus of genomic and evolutionary studies (Figure 1) (Fedoroff, 2012; Zhao et al., 2018; Stitzer et al., 2019; Wang et al., 2020; Noshay et al., 2020; Sharma and Peterson, 2022).
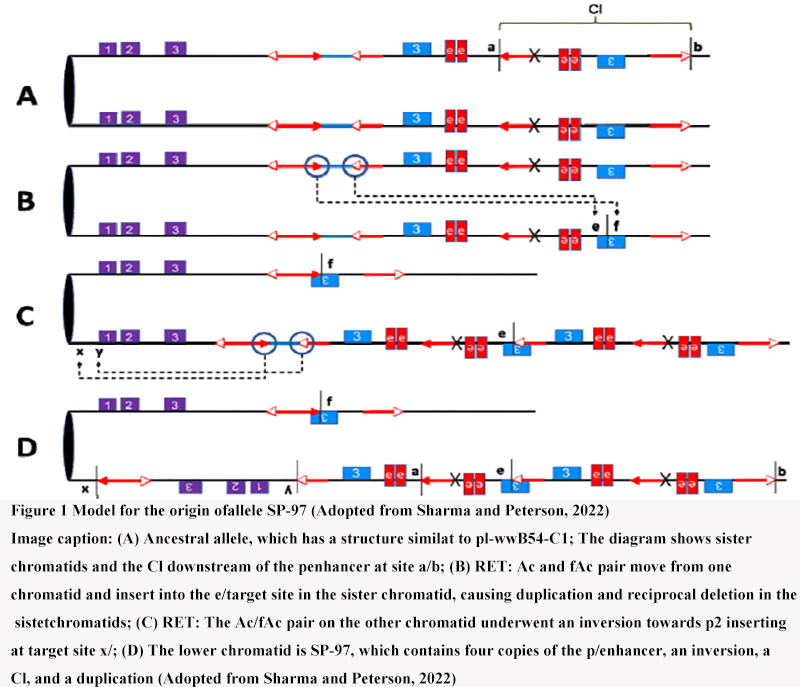 Figure 1 Model for the origin ofallele SP-97 (Adopted from Sharma and Peterson, 2022) Image caption: (A) Ancestral allele, which has a structure similat to pl-wwB54-C1; The diagram shows sister chromatids and the Cl downstream of the penhancer at site a/b; (B) RET: Ac and fAc pair move from one chromatid and insert into the e/target site in the sister chromatid, causing duplication and reciprocal deletion in the sistetchromatids; (C) RET: The Ac/fAc pair on the other chromatid underwent an inversion towards p2 inserting at target site x/; (D) The lower chromatid is SP-97, which contains four copies of the p/enhancer, an inversion, a Cl, and a duplication (Adopted from Sharma and Peterson, 2022) |
.png){kind=link}
3 Mechanisms of Transposition
3.1 How TEs move within the genome
Transposable elements (TEs) are dynamic components of the genome that can move from one location to another, a process known as transposition. This movement can occur through two primary mechanisms: "cut-and-paste" and "copy-and-paste." The "cut-and-paste" mechanism involves DNA transposons, which excise themselves from one genomic location and integrate into another. This process is facilitated by the enzyme transposase, which recognizes specific sequences at the ends of the transposon, cuts the DNA, and inserts the transposon into a new site (Fedoroff, 2012; Bennetzen and Wang, 2014; Romano and Fanti, 2022).
The "copy-and-paste" mechanism is characteristic of retrotransposons, which move via an RNA intermediate. Retrotransposons are transcribed into RNA, which is then reverse-transcribed into DNA by the enzyme reverse transcriptase. The newly synthesized DNA is then integrated into a new genomic location. This mechanism not only increases the number of TEs within the genome but also contributes to genome expansion (Fedoroff, 2012; Bennetzen and Wang, 2014; Klein and O’Neill, 2018).
3.2 Enzymatic processes involved
The movement of TEs within the genome is facilitated by specific enzymes, primarily transposases and reverse transcriptases. Transposases are crucial for the "cut-and-paste" mechanism of DNA transposons. These enzymes recognize terminal inverted repeats (TIRs) at the ends of the transposon, excise the transposon from its original location, and integrate it into a new site. This process is highly specific and requires precise recognition of the TIRs (Fedoroff, 2012; Bennetzen and Wang, 2014; Romano and Fanti, 2022).
Reverse transcriptases are essential for the "copy-and-paste" mechanism of retrotransposons. These enzymes transcribe the RNA intermediate back into DNA, which is then integrated into the genome. The integration process is often facilitated by integrase, another enzyme that helps insert the newly synthesized DNA into the host genome (Fedoroff, 2012). The activity of these enzymes is tightly regulated to prevent uncontrolled transposition, which can lead to genomic instability (Bennetzen and Wang, 2014; Klein and O’Neill, 2018).
3.3 Regulatory mechanisms controlling TE activity
The activity of TEs is tightly regulated by the host genome to prevent potential deleterious effects such as gene disruption, chromosomal rearrangements, and genome instability. One of the primary regulatory mechanisms is epigenetic modification, including DNA methylation and histone modification. These modifications can silence TEs by altering the chromatin structure, making it less accessible for transcription and transposition (Figure 2) (Friedli and Trono, 2015; Vicient and Casacuberta, 2017; Schrader and Schmitz, 2018).
 Figure 2 TEs as ‘molecular parasites’ or ‘functional symbionts’ (Adopted from Romano and Fanti, 2022) Image caption: As ‘molecular parasites’ TEs can produce a variety of detrimental effects on the host genome. (a) The insertion of TEs within coding exons can cause frame shift mutations disrupting protein sequence and function. (b) TEs can cause genomic instability being the substrate for chromosome rearrangements, such as duplications, deletions, inversions and translocations. (c) The insertions of TEs in regulatory stretches such as in 50 or 30 regions or introns can cause epigenetic modifications resulting in inappropriate activation or repression of gene expression. The co-option of TEs by the host genome may generate new regulatory signals or coding sequences. This process is referred to as ‘molecular domestication’. (d) TEs may contribute new enhancer sequences for transcription factors (grey circle) changing the spatial/temporal regulation of gene expression. (e) After the loss of telomerase, retrotransposons can actively participate in the maintenance of telomeres. Three non-LTR families, HeT-A, TAHRE, and TART form a head-to-tail array. They express Gag and Reverse Transcriptase proteins that are necessary for the elongation of telomeres. (f) TEs can contribute to the maintenance of genome architecture by providing binding sites for the CTCF protein that is responsible for establishing “topologically associated domains” (TADs) (Adopted from Romano and Fanti, 2022) |
.png){kind=link}
4 TEs and Genetic Diversity
4.1 Creation of genetic variation through TE insertions and excisions
Transposable elements (TEs) are a significant source of genetic variation in maize, contributing to both the structure and function of the genome. TEs can create genetic diversity through their ability to insert and excise themselves within the genome. This process can lead to mutations, gene duplications, and the creation of new regulatory elements, all of which can have profound effects on gene expression and phenotype.
The maize genome is particularly rich in TEs, with more than 85% of its sequence attributed to past transposition events (Stitzer et al., 2019). This high TE content has been shown to play a crucial role in shaping the genome's structure and function. For instance, TEs can insert themselves into or near genes, potentially disrupting gene function or altering gene expression patterns. These insertions can be beneficial, neutral, or deleterious, depending on their location and the genes they affect (Figure 3) (Qiu et al., 2021).
.png) Figure 3 Relationship between TE frequency and relative position to the nearest gene (Adopted from Qiu et al., 2021) Image caption: Helitrons (A), TIRs (B), and LTRs (C) were categorized using the following hierarchy: gene completely within the TE, TE completely within the 5’ UTR, completely within the 3’ UTR, completely within an exon, completely within an intron, completely encompassed by a gene, 0–1 kb upstream of a gene, 1–5 kb upstream of a gene, 5–10 kb upstream of a gene, 0–1 kb downstream of a gene, 1–5 kb downstream of a gene, 5–10 kb downstream of a gene, intergenic (not shown in figure). Full colored bars (left bars) contain frequencies for TEs in each genomic region and transparent bars (right bars) contain frequencies for genome-wide background rates based on 100 bootstrap iterations. (D) Proportion of LTRs with low LTR similarity (LTR similarity <95%), moderate LTR similarity (LTR similarity between 95 and 99%), and high LTR similarity (LTR similarity >99%) in each gene proximity category (Adopted from Qiu et al., 2021) |
{kind=link}
TEs can also excise themselves from the genome, a process that can restore gene function if the TE was previously disruptive. However, excision is not always precise and can leave behind small insertions or deletions, further contributing to genetic variation. The dynamic nature of TE insertions and excisions thus provides a continuous source of genetic diversity, which can be acted upon by natural selection (Vicient and Casacuberta, 2017).
TEs can influence the epigenetic landscape of the genome. For example, the insertion of a TE can attract epigenetic modifications such as DNA methylation, which can spread to nearby genes and alter their expression. This epigenetic influence can be heritable, adding another layer of complexity to the genetic variation generated by TEs (Su et al., 2019).
4.2 Examples of phenotypic changes in maize due to TE activity
The activity of TEs in maize has been linked to several notable phenotypic changes, illustrating their role in driving genetic diversity and evolution. One classic example is the role of TEs in the pigmentation of maize kernels. The insertion of a TE in the regulatory region of the C1 gene, which controls anthocyanin biosynthesis, can lead to variegated kernel coloration. When the TE excises, it can restore the gene's function, resulting in patches of pigmented tissue (Schrader and Schmitz, 2018).
Another example is the impact of TEs on flowering time in maize. The insertion of a TE in the promoter region of the ZmCCT9 gene has been associated with delayed flowering. This insertion disrupts the normal expression of the gene, leading to changes in the plant's developmental timing. Such variations in flowering time can have significant implications for adaptation to different environmental conditions (Pimpinelli and Piacentini, 2020).
TEs have also been implicated in the evolution of disease resistance in maize. The insertion of a TE in the promoter region of the Rp1-D gene, which confers resistance to rust pathogens, can enhance the gene's expression and provide increased resistance. This example highlights how TEs can contribute to the rapid evolution of traits that are critical for plant survival and fitness (Wells and Feschotte, 2020).
Furthermore, TEs can generate novel regulatory elements that drive the expression of nearby genes in new patterns. For instance, the insertion of a TE upstream of the tb1 gene, which controls plant architecture, has been shown to create a new enhancer element. This enhancer drives higher expression of tb1, leading to the characteristic upright growth habit of domesticated maize compared to its wild ancestor, teosinte (Fedoroff, 2012).
5 Transposable Elements and Genome Evolution
5.1 Impact on genome structure and function
Transposable elements (TEs) are significant contributors to the structural and functional dynamics of genomes. They can cause chromosome rearrangements, influence genome size, and affect gene content and order. TEs are known to drive centromere function and other aspects of nuclear biology, often acting as agents of genomic novelty (Bennetzen and Wang, 2014). In plants, TEs can lead to drastic changes in genome size and provide new coding and regulatory sequences, which are particularly impactful following polyploidization events. These elements can induce bursts of transposition, leading to increased rates of gene mutations and changes in gene regulation due to their insertion near or within genes (Vicient and Casacuberta, 2017). Additionally, TEs can cause inter-element recombination, resulting in large-scale genome rearrangements and reductions in genome size, a process known as diploidization (Vicient and Casacuberta, 2017).
5.2 Role in gene regulation and expression
TEs play a crucial role in the regulation of gene expression. They can provide cis-regulatory sequences that influence the expression of nearby genes. These sequences can act as enhancers, promoters, silencers, and boundary elements, thereby facilitating changes in gene regulatory networks (Sundaram and Wysocka, 2020). TEs can also produce regulatory RNAs, such as miRNAs and long non-coding RNAs (lncRNAs), which are involved in post-transcriptional regulation (Ali et al., 2020). In plants, TEs can influence gene expression through various mechanisms, including the provision of alternative promoters and the alteration of chromatin modifications near genes (Hirsch and Springer, 2017). The production of short interfering RNAs (siRNAs) by TEs is another mechanism through which they regulate gene expression, particularly in polyploid plants where TEs from one parent can affect the TEs of the other parent, leading to complex epigenetic regulation (Gill et al., 2021).
5.3 Contribution to the evolution of new gene functions
TEs are not merely passive elements within the genome; they actively contribute to the evolution of new gene functions. They can be co-opted by the host genome to create new genes or modify existing ones. This co-option can lead to the development of novel gene functions and regulatory mechanisms (Bennetzen and Wang, 2014). TEs have been shown to contain functional binding sites for transcription factors, contributing to the evolution of gene regulatory networks (Sundaram et al., 2014). In mammals, TEs have been implicated in morphological evolution by altering gene regulatory networks and genome architecture (Nishihara, 2019). The ability of TEs to provide new regulatory sequences and influence gene expression patterns makes them potent drivers of evolutionary innovation (Sundaram and Wysocka, 2020). The waves of TE invasions over evolutionary time have catalyzed the development of complex gene-regulatory networks, highlighting their role in the adaptive evolution of genomes (Chuong et al., 2016).
TEs are pivotal in shaping genome structure and function, regulating gene expression, and driving the evolution of new gene functions. Their ability to induce genomic changes and provide new regulatory elements underscores their importance in the evolutionary dynamics of genomes.
6 Transposable Elements as Agents of Evolutionary Innovation
6.1 Examples of adaptive traits in maize linked to TEs
Transposable elements (TEs) have been instrumental in shaping the genetic landscape of maize, contributing to its adaptive traits and overall genetic diversity. One notable example is the role of TEs in generating allelic diversity and structural variation, which can lead to significant phenotypic changes. For instance, TIR elements, a type of Class II DNA TEs, have been shown to influence gene expression and genome structure in maize. The development of new methods for TIR element detection has revealed a much larger presence of these elements in the maize genome than previously thought, highlighting their potential impact on adaptive traits (Su et al., 2019).
Moreover, TEs can facilitate adaptive responses to environmental challenges by increasing genome plasticity. This plasticity allows for rapid genetic changes that can be beneficial under stress conditions, such as drought or pest resistance. The ability of TEs to induce such changes is particularly relevant in species like maize, which frequently encounter novel environments (Schrader and Schmitz, 2018). Additionally, TEs can contribute to the formation of new regulatory sequences and genes through processes like exaptation, where TE sequences are co-opted for new functions beneficial to the host (Etchegaray et al., 2021).
6.2 Potential for TEs to drive speciation
The role of TEs in speciation is a burgeoning area of research, with evidence suggesting that these elements can contribute to reproductive iso lation and the emergence of new species. TEs can alter regulatory networks and gene expression, leading to significant genomic rearrangements that may result in hybrid defects. These defects can act as barriers to gene flow between nascent species, thereby facilitating speciation (Serrato-Capuchina and Matute, 2018).
In maize, the high activity and diversity of TEs create a dynamic genomic environment that can drive speciation. The interplay between TEs and the host genome can lead to the development of unique genetic traits that distinguish one population from another. For example, the insertion of TEs near genes involved in reproductive processes can result in changes that prevent interbreeding between populations, thus promoting speciation (Stitzer et al, 2019).
Furthermore, TEs can contribute to the evolution of lineage-specific traits by providing raw genetic material for innovation. In vertebrates, for instance, TEs have been implicated in the evolution of key innovations such as adaptive immunity and complex brain structures (Warren et al., 2015). Similarly, in maize, TEs can drive the evolution of traits that are crucial for survival and reproduction in specific environments, thereby contributing to the diversification of the species.
TEs are not merely passive elements within the genome but active players in the evolutionary process. Their ability to induce genetic variability and influence gene expression makes them powerful agents of evolutionary innovation. In maize, TEs have been linked to adaptive traits and have the potential to drive speciation by creating reproductive barriers and facilitating the evolution of unique genetic traits. Understanding the role of TEs in these processes is crucial for comprehending the broader mechanisms of evolution and genetic diversity in plants.
7 TEs and Genomic Stability
7.1 Negative impacts of TE activity
Transposable elements (TEs) are known to significantly impact genomic stability, often leading to deleterious consequences. The mobilization of TEs can cause mutations, chromosomal rearrangements, and gene disruptions, which can result in diseases and other dysfunctions. For instance, TEs can induce harmful mutations by inserting themselves into essential genes or regulatory regions, thereby disrupting normal gene function and potentially leading to lethality (Klein and O’Neill, 2018; Schrader and Schmitz, 2018; Romano and Fanti, 2022). In the fungal wheat pathogen Zymoseptoria tritici, TE activity has been shown to destabilize genome integrity, although it can also confer adaptive variation in pathogenicity or resistance traits (Oggenfuss and Croll, 2023). Moreover, the proliferation of TEs can lead to genome instability by creating novel chromosome rearrangements and impacting gene expression, which can result in disease in some cases and species-specific diversity in others (Klein and O’Neill, 2018). In Drosophila species, TE mobilization poses a constant threat to genome integrity, necessitating robust defensive mechanisms to suppress their activity (Wei et al., 2022). The negative association between TE insertion frequency and the expression of nearby genes suggests that TEs tend to insert near lowly expressed, nonessential genes to minimize fitness impacts (Wei et al., 2022).
7.2 Mechanisms of TE suppression and genomic defense
To counteract the deleterious effects of TEs, organisms have evolved several defense mechanisms that act at various levels, including epigenetic, transcriptional, and post-transcriptional levels (Castanera et al., 2016; Platt et al., 2018; Romano and Fanti, 2022). One of the primary defense mechanisms involves RNA-mediated silencing, which plays a crucial role in limiting TE proliferation. This system uses small RNAs to target and silence TEs, thereby preventing their mobilization and subsequent genomic instability (Blumenstiel, 2011).
Epigenetic mechanisms, such as DNA methylation and histone modifications, are also vital in maintaining TE silencing. For example, in fungi, active cytosine methylation machinery is associated with TE-mediated gene silencing, suggesting that epigenetic defenses are crucial in controlling TE proliferation (Castanera et al., 2016). Additionally, the stress-dependent incapacitation of these defense mechanisms can facilitate adaptive responses to environmental challenges, highlighting the dynamic interplay between TEs and host genomes (Schrader and Schmitz, 2018).
The evolutionary arms race between TEs and host genomes is characterized by TEs evolving to escape suppression and host genomes developing compensatory changes to re-establish control. This ongoing conflict drives the rapid evolution of TEs and the recurrent positive selection of genes involved in host defense (Platt et al., 2018; Wei et al., 2022). In plants, epigenetic regulation has been transformed from a process to silence invading TEs and viruses into a key strategy for regulating plant genes, demonstrating the co-option of TE components by the host to create new genes or modify gene regulation (Bennetzen and Wang, 2014). The suppression of TEs is a complex and multifaceted process involving various genomic defense mechanisms that work together to maintain genomic stability and prevent the deleterious effects of TE activity.
8 Comparative Analysis with Other Species
8.1 Comparison of TE activity and impact in Zea versus other plants and vertebrates
Transposable elements (TEs) are ubiquitous across eukaryotic genomes, playing significant roles in genetic diversity and evolution. In Zea (maize), TEs constitute a substantial portion of the genome, contributing to its large size and complexity. This is comparable to other plant species where TEs also play a crucial role in genome evolution. For instance, in Arabidopsis lyrata, selfing populations exhibit higher TE copy numbers and allele frequencies, suggesting that TEs can proliferate more in certain mating systems (Bonchev and Willi, 2018). Similarly, in polyploid plants, TEs are major drivers of genome size changes and new coding/regulatory sequences, often activated by polyploidization events (Vicient and Casacuberta, 2017).
In vertebrates, the impact of TEs varies significantly across lineages. For example, in teleost fish, TEs contribute more to genome size compared to mammals, with DNA transposons being more prevalent in fish genomes (Chalopin et al., 2015). In contrast, mammalian genomes are predominantly shaped by non-long terminal repeat (non-LTR) retrotransposons, with older TE sequences persisting over time (Chalopin et al., 2015). This difference in TE composition and activity between vertebrates and plants highlights the diverse evolutionary pressures and mechanisms influencing TE dynamics.
Moreover, the diversity of TEs in vertebrates is notable, with some lineages like ray-finned fishes and amphibians exhibiting higher TE diversity compared to birds and mammals (Sotero-Caio et al., 2017). This is akin to the high diversity and dynamic nature of TEs observed in land plants, where genetic and epigenetic factors modulate TE activity and distribution (Mhiri et al., 2022). The evolutionary impact of TEs in vertebrates includes their role in chromosome structure, gene regulation, and the formation of non-coding RNAs and protein-coding genes, driving lineage-specific innovations.
8.2 Insights from cross-species studies
Cross-species studies provide valuable insights into the evolutionary roles of TEs. In Zea, TEs are not only a source of genetic variation but also influence gene expression and genome plasticity. This is similar to the role of TEs in fungal phytopathogens, where they contribute to genome diversification, pathogenicity, and adaptive evolution (Razali et al., 2019). The ability of TEs to alter gene expression by modifying cis-regulatory elements or recruiting epigenetic control is a common theme across different species, underscoring their importance in adaptive responses to environmental challenges (Schrader and Schmitz, 2018; Razali et al., 2019).
In vertebrates, comparative analyses of TE content across multiple genomes reveal significant variations in TE abundance and activity. For instance, the zebrafish genome has a high TE content (55%), while the pufferfish genome has a much lower TE content (6%) (Chalopin et al., 2015). These differences are indicative of lineage-specific TE dynamics and evolutionary histories. The loss of major TE families in sarcopterygians, particularly in birds and mammals, suggests a reduction in TE diversity over time, possibly due to stronger selective pressures against TE proliferation in these lineages (Chalopin et al., 2015).
Furthermore, the role of TEs in speciation is an emerging area of interest. TEs can cause reproductive isolation by inducing hybrid defects, thereby contributing to the formation of new species (Warren et al., 2015). This phenomenon is observed across diverse taxa, highlighting the potential of TEs to drive speciation through genetic incompatibilities and regulatory network alterations.
Cross-species studies emphasize the multifaceted roles of TEs in genome evolution. In plants, TEs are major players in genome size variation and gene regulation, while in vertebrates, they contribute to genomic diversity and lineage-specific innovations. The comparative analysis of TE activity and impact across different species enhances our understanding of their evolutionary significance and the mechanisms underlying their dynamic behavior.
9 Future Research Directions
9.1 Technological advancements in TE research
The advent of long-read sequencing technologies, such as those provided by Oxford Nanopore and PacBio, has revolutionized the study of transposable elements (TEs) in plant genomes, including maize (Zea mays). These technologies enable the generation of longer sequence reads, which significantly improve the accuracy of TE annotation and the identification of transcriptionally active elements. For instance, long-read cDNA sequencing has been employed to create a transcript-based annotation of TEs in Arabidopsis thaliana and maize, revealing new insights into TE transcription start sites, polyadenylation sites, and splicing patterns (Benoit, 2020; Panda et al., 2020). This approach reduces the bioinformatic complexity associated with repetitive TEs and enhances the resolution of TE mapping, which is often hampered by the short read lengths of second-generation sequencing technologies (Benoit, 2020).
Moreover, long-read sequencing has facilitated the identification of previously unannotated TEs and the discovery of new TE families, as demonstrated by the TIR-Learner method, which combines homology-based and de novo machine-learning approaches to improve TIR element annotation in the maize genome (Su et al., 2019). This method has revealed a much larger proportion of TIR elements than previously recognized, highlighting the potential of advanced sequencing technologies to uncover hidden genomic diversity.
Future research should focus on further integrating long-read sequencing with other genomic technologies, such as chromatin immunoprecipitation sequencing (ChIP-seq) and Hi-C, to explore the regulatory roles of TEs in genome architecture and gene expression. Additionally, the development of more sophisticated bioinformatic tools to handle the vast amounts of data generated by long-read sequencing will be crucial for advancing our understanding of TE dynamics and their impact on genetic diversity and evolution.
9.2 Unresolved questions and potential research areas
Despite significant progress in TE research, several unresolved questions and potential research areas remain. One major question is the precise role of TEs in speciation and reproductive isolation. While TEs have been implicated in hybrid defects that may prevent gene flow between nascent species, their involvement in other barriers to gene flow, such as prezygotic isolation, is still not well understood (Serrato-Capuchina and Matute, 2018). Systematic studies are needed to elucidate the mechanisms by which TEs contribute to speciation and to identify the specific TE families involved in these processes.
Another important area of research is the impact of TEs on genome stability and their potential role in health and disease. TEs can cause genomic instability through gene disruption, inversions, deletions, and duplications, but they also play roles in gene regulation and epigenetic control (Bhat et al., 2022). Understanding the balance between the deleterious and beneficial effects of TEs on genome stability is crucial for developing therapeutic applications that harness TEs for gene editing and disease treatment.
The adaptive roles of TEs in genome plasticity and pathogenicity, particularly in fungal phytopathogens, represent a promising research avenue. TEs can drive genome diversification and the emergence of novel pathogenicity factors, but the exact mechanisms behind these processes remain poorly understood (Razali et al., 2019). Investigating the interactions between TEs and host genomes in different environmental contexts will provide insights into the evolutionary strategies of both TEs and their hosts.
The diversity and evolutionary dynamics of TEs across different taxa, including vertebrates and plants, warrant further exploration. Comparative analyses of TE content and activity in various species have revealed significant variations and lineage-specific patterns, but the evolutionary origins and relationships of different TE families are still not fully resolved (Chalopin et al., 2015; Sotero-Caio et al., 2017). Expanding these comparative studies to include a broader range of species and integrating them with functional genomics approaches will enhance our understanding of the evolutionary impact of TEs on genome diversity and adaptation. Future research directions in TE studies should leverage technological advancements, address unresolved questions, and explore new potential research areas to deepen our understanding of the roles of TEs in genetic diversity and evolution.
10 Concluding Remarks
Transposable elements (TEs) are integral components of the maize (Zea mays) genome, playing a crucial role in genetic diversity and evolution. This section summarizes the key points discussed and underscores the overarching importance of TEs in the genetic landscape of Zea. TEs are ubiquitous genetic elements capable of moving within the genome, significantly impacting genetic variability and evolutionary processes. They can induce mutations, which may be deleterious, but also contribute to genetic diversity, enabling populations to adapt to environmental changes. In maize, TEs constitute a substantial portion of the genome, with Class II DNA TEs, particularly TIR elements, being prominent. These elements generate allelic diversity, induce structural variations, and regulate gene expression, thereby influencing genome evolution.
The evolutionary impact of TEs extends beyond mere genetic variability. They are involved in the regulation of gene expression, formation of non-coding RNAs, and the evolution of protein-coding genes. This multifaceted role underscores their importance in the adaptive evolution of species. In maize, the coevolution of TEs with RNA modifications, such as N6-methyladenosine, further highlights their role in genome plasticity and response to environmental stress.
TEs also contribute to the structural and functional diversity of genomes. TEs can lead to the formation of new genes and regulatory networks, which are essential for the evolutionary success of species. In maize, the identification of new TIR elements through advanced annotation methods has revealed a higher than previously estimated TE content, emphasizing their significant role in genome diversity. TEs are not merely parasitic elements but are symbionts of the genome, driving genetic diversity and evolution. Their ability to induce genetic variability, regulate gene expression, and contribute to genome architecture makes them indispensable in the study of maize genetics. The dynamic interplay between TEs and the host genome underscores their pivotal role in the evolutionary processes that shape the genetic landscape of Zea.
Acknowledgments
The author extends sincere thanks to two anonymous peer reviewers for their feedback on the manuscript.
Conflict of Interest Disclosure
The author affirms that this research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Ali A., Han K., and Liang P., 2020, Role of transposable elements in gene regulation in the human genome, Life, 11(2): 118.
https://doi.org/10.3390/life11020118
PMid:33557056 PMCid:PMC7913837
Bennetzen, J., and Wang, H., 2014, The contributions of transposable elements to the structure, function, and evolution of plant genomes, Annual Review of Plant biology, 65: 505-530.
https://doi.org/10.1146/annurev-arplant-050213-035811
PMid:24579996
Benoit M., 2020, In the transcripts: long-read transcriptomics enables a novel type of transposable element annotation in plants, The Plant Cell, 32(9): 2661-2662.
https://doi.org/10.1105/tpc.20.00523
PMid:32665309 PMCid:PMC7474303
Bhat A., Ghatage T., Bhan S., Lahane G., Dhar A., Kumar R., Pandita R., Bhat K.M., Ramos K.S., and Pandita T., 2022, Role of transposable elements in genome stability: implications for health and disease, International Journal of Molecular Sciences, 23(14): 7802.
https://doi.org/10.3390/ijms23147802
Blumenstiel J., 2011, Evolutionary dynamics of transposable elements in a small RNA world, Trends in Genetics, 27(1): 23-31.
https://doi.org/10.1016/j.tig.2010.10.003
Bonchev G., and Willi Y., 2018, Accumulation of transposable elements in selfing populations of Arabidopsis lyrata supports the ectopic recombination model of transposon evolution, The New Phytologist, 219(2): 767-778.
https://doi.org/10.1111/nph.15201
PMid:29757461
Castanera R., López-Varas L., Borgognone A., LaButti K., Lapidus A., Schmutz J., Grimwood J., Pérez G., Pisabarro A., Grigoriev I., Stajich J., and Ramírez L., 2016, Transposable elements versus the fungal genome: impact on whole-genome architecture and transcriptional profiles, PLoS Genetics, 12(6): e1006108.
https://doi.org/10.1371/journal.pgen.1006108
PMid:27294409 PMCid:PMC4905642
Chalopin D., Naville M., Plard F., Galiana D., and Volff J., 2015, Comparative analysis of transposable elements highlights mobilome diversity and evolution in vertebrates, Genome Biology and Evolution, 7: 567-580.
https://doi.org/10.1093/gbe/evv005
PMid:25577199 PMCid:PMC4350176
Chénais B., Caruso A., Hiard S., and Casse N., 2012, The impact of transposable elements on eukaryotic genomes: from genome size increase to genetic adaptation to stressful environments, Gene, 509(1): 7-15.
https://doi.org/10.1016/j.gene.2012.07.042
PMid:22921893
Chuong, E., Elde, N., and Feschotte, C., 2016, Regulatory activities of transposable elements: from conflicts to benefits, Nature Reviews Genetics, 18: 71-86.
https://doi.org/10.1038/nrg.2016.139.
PMid:27867194 PMCid:PMC5498291
Drongitis D., Aniello F., Fucci L., and Donizetti A., 2019, Roles of transposable elements in the different layers of gene expression regulation, International Journal of Molecular Sciences, 20(22): 5755.
https://doi.org/10.3390/ijms20225755
PMid:31731828 PMCid:PMC6888579
Etchegaray E., Naville M., Volff J., and Haftek-Terreau Z., 2021, Transposable element-derived sequences in vertebrate development, Mobile DNA, 12: 1-24.
https://doi.org/10.1186/s13100-020-00229-5
Fedoroff N., 2012, Transposable elements, epigenetics, and genome evolution, Science, 338: 758-767.
https://doi.org/10.1126/science.338.6108.758
PMid:23145453
Friedli M., and Trono D., 2015, The developmental control of transposable elements and the evolution of higher species, Annual Review of Cell and Developmental Biology, 31: 429-451.
https://doi.org/10.1146/annurev-cellbio-100814-125514
PMid:26393776
Gill R., Scossa F., King G., Golicz A., Tong C., Snowdon R., Fernie A., and Liu S., 2021, On the role of transposable elements in the regulation of gene expression and subgenomic interactions in crop genomes, Critical Reviews in Plant Sciences, 40: 157-189.
https://doi.org/10.1080/07352689.2021.1920731
Hirsch C., and Springer N., 2017, Transposable element influences on gene expression in plants, biochimica et biophysica acta, Gene Regulatory Mechanisms, 1860(1): 157-165.
https://doi.org/10.1016/j.bbagrm.2016.05.010
PMid:27235540
Klein S., and O'Neill R., 2018, Transposable elements: genome innovation, chromosome diversity, and centromere conflict, Chromosome Research, 26: 5-23.
https://doi.org/10.1007/s10577-017-9569-5.
PMid:29332159 PMCid:PMC5857280
Mhiri C., Borges F., and Grandbastien M., 2022, Specificities and dynamics of transposable elements in land plants, Biology, 11(4): 488.
PMid:35453688 PMCid:PMC9033089
Nishihara H., 2019, Transposable elements as genetic accelerators of evolution: contribution to genome size, gene regulatory network rewiring and morphological innovation, Genes and Genetic Systems, 94(6): 269-281.
https://doi.org/10.1266/ggs.19-00029
PMid:31932541
Noshay J., Marand A., Anderson S., Zhou P., Guerra M., Lu Z., O'Connor C., Crisp P., Hirsch C., Schmitz R., and Springer N., 2020, Assessing the regulatory potential of transposable elements using chromatin accessibility profiles of maize transposons., Genetics, 217(1): 1-13.
https://doi.org/10.1093/genetics/iyaa003
PMid:33683350 PMCid:PMC8552484
Oggenfuss U., and Croll D., 2023, Recent transposable element bursts are associated with the proximity to genes in a fungal plant pathogen, PLOS Pathogens, 19(2): e1011130.
https://doi.org/10.1371/journal.ppat.1011130
PMid:36787337 PMCid:PMC9970103
Panda K., Slotkin R., and Slotkin R., 2020, Long-read cDNA sequencing enables a "gene-like" transcript annotation of transposable elements, Plant Cell, 32: 2687-2698.
https://doi.org/10.1105/tpc.20.00115
PMid:32647069 PMCid:PMC7474280
Pimpinelli S., and Piacentini L., 2020, Environmental change and the evolution of genomes: transposable elements as translators of phenotypic plasticity into genotypic variability, Functional Ecology, 34: 428-441.
https://doi.org/10.1111/1365-2435.13497
Platt R., Vandewege M., and Ray D., 2018, Mammalian transposable elements and their impacts on genome evolution, Chromosome Research, 26: 25-43.
https://doi.org/10.1007/s10577-017-9570-z.
PMid:29392473 PMCid:PMC5857283
Qiu Y., O'Connor C., Coletta R., Renk J., Monnahan P., Noshay J., Liang Z., Gilbert A., Anderson S., McGaugh S., Springer N., and Hirsch C., 2021, Whole-genome variation of transposable element insertions in a maize diversity panel, G3: Genes|Genomes|Genetics, 11(10): p.jkab238.
https://doi.org/10.1093/g3journal/jkab238.
PMid:34568911 PMCid:PMC8473971
Razali N., Cheah B., and Nadarajah K., 2019, Transposable elements adaptive role in genome plasticity, pathogenicity and evolution in fungal phytopathogens, International Journal of Molecular Sciences, 20(14): 3597.
https://doi.org/10.3390/ijms20143597
PMid:31340492 PMCid:PMC6679389
Rebollo R., Romanish M., and Mager D., 2012, Transposable elements: an abundant and natural source of regulatory sequences for host genes, Annual Review of Genetics, 46: 21-42.
https://doi.org/10.1146/annurev-genet-110711-155621
PMid:22905872
Romano N., and Fanti L., 2022, Transposable elements: major players in shaping genomic and evolutionary patterns, Cells, 11(6): 1048.
https://doi.org/10.3390/cells11061048.
PMid:35326499 PMCid:PMC8947103
Schrader L., and Schmitz J., 2018, The impact of transposable elements in adaptive evolution, Molecular Ecology, 28: 1537-1549.
https://doi.org/10.1111/mec.14794.
PMid:30003608
Serrato-Capuchina A., and Matute D., 2018, The role of transposable elements in speciation, Genes, 9(5): 254.
https://doi.org/10.3390/genes9050254
Sharma S., and Peterson T., 2022, Complex chromosomal rearrangements induced by transposons in maize, Genetics, 223(2): iyac124.
https://doi.org/10.1093/genetics/iyac124.
PMid:36111993 PMCid:PMC9910405
Sotero-Caio C., Platt R., Suh A., and Ray D., 2017, Evolution and diversity of transposable elements in vertebrate genomes, Genome Biology and Evolution, 9: 161-177.
https://doi.org/10.1093/gbe/evw264
PMid:28158585 PMCid:PMC5381603
Stitzer M., Anderson S., Springer N., and Ross-Ibarra J., 2019, The genomic ecosystem of transposable elements in maize, PLoS Genetics, 17(10): e1009768.
https://doi.org/10.1101/559922.
Su W., Gu X., and Peterson T., 2019, TIR-Learner, a new ensemble method for TIR transposable element annotation, provides evidence for abundant new transposable elements in the maize genome, Molecular Plant, 12(3): 447-460.
https://doi.org/10.1016/j.molp.2019.02.008
PMid:30802553
Sundaram V., and Wysocka J., 2020, Transposable elements as a potent source of diverse cis-regulatory sequences in mammalian genomes, Philosophical Transactions of the Royal Society B: Biological Sciences, 375(1795): 20190347.
https://doi.org/10.1098/rstb.2019.0347
PMid:32075564 PMCid:PMC7061989
Sundaram V., Cheng Y., Ma Z., Li D., Xing X., Edge P., Snyder M., and Wang T., 2014, Widespread contribution of transposable elements to the innovation of gene regulatory networks, Genome Research, 24: 1963-1976.
https://doi.org/10.1101/gr.168872.113
PMid:25319995 PMCid:PMC4248313
Vicient C., and Casacuberta J., 2017, Impact of transposable elements on polyploid plant genomes, Annals of Botany, 120(2): 195-207.
https://doi.org/10.1093/aob/mcx078.
PMid:28854566 PMCid:PMC5737689
Wang D., Zhang J., Zuo T., Zhao M., Lisch D., and Peterson T., 2020, Small RNA-mediated de novo silencing of Ac/Ds transposons is initiated by alternative transposition in maize, Genetics, 215(2): 393-406.
https://doi.org/10.1534/genetics.120.303264
PMid:32317287 PMCid:PMC7268987
Warren I., Naville M., Chalopin D., Levin P., Berger C., Galiana D., and Volff J., 2015, Evolutionary impact of transposable elements on genomic diversity and lineage-specific innovation in vertebrates, Chromosome Research, 23: 505-531.
https://doi.org/10.1007/s10577-015-9493-5
PMid:26395902
Wei K., Mai D., Chatla K., and Bachtrog D., 2022, Dynamics and impacts of transposable element proliferation in the drosophila nasuta species group radiation, Molecular Biology and Evolution, 39(5): msac080.
https://doi.org/10.1093/molbev/msac080
PMid:35485457 PMCid:PMC9075770
Wells J., and Feschotte C., 2020, A field guide to eukaryotic transposable elements, Annual Review of Genetics, 54(1): 539-561.
https://doi.org/10.1146/annurev-genet-040620-022145
PMid:32955944 PMCid:PMC8293684
Zhao H., Zhang W., Chen L., Wang L., Marand A., Wu Y., and Jiang J., 2018, Proliferation of regulatory DNA elements derived from transposable elements in the maize genome, Plant Physiology, 176(4): 2789-2803.
https://doi.org/10.1104/pp.17.01467
PMid:29463772 PMCid:PMC5884613




. PDF(784KB)
. FPDF(win)
. FPDF(mac)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Shaomin Yang
Related articles
. Transposable elements
. Genetic diversity
. Evolution
. Zea
. Epigenetic regulation
Tools
. Email to a friend
. Post a comment